Author:
A group of researches: G. Lithgow, S. Murakami, T. Johnson; L. Guarente, C. Kenyon,
History:
This theory is as it were the theory of
 Example:
Example:
Under stress, the organism enters the «waiting mode». In this mode, the organism saves its resources suppressing biosynthesis of structural proteins, switching the household genes off, holding the growth and reproduction up. Besides, the organism delays the entry into the reproduction cycle and increases its stress resistance (it means that the organism activates its antioxidant systems, induces
Description of the Theory:

According to this theory, the program of longevity might appear in evolution to overcome
 Molecular and cellular mechanisms of stress resistance turning on in the «maintenance mode» can afterwards help to overcome another sorts of stress (e.g. negative effects of radiation). The same mechanisms withstand the
Molecular and cellular mechanisms of stress resistance turning on in the «maintenance mode» can afterwards help to overcome another sorts of stress (e.g. negative effects of radiation). The same mechanisms withstand the
Mutations are currently known which increase the life span in experimental animal. Individuals with such mutations get into the «maintenance mode» irrespective of the environmental conditions.
Additions and Сriticism:
 While the mutation accumulation and the disposable soma theories of aging rest upon permanent pressure of the environmental conditions (predators, diseases), the program of longevity could arise in evolution for overcoming the
While the mutation accumulation and the disposable soma theories of aging rest upon permanent pressure of the environmental conditions (predators, diseases), the program of longevity could arise in evolution for overcoming the
The programmed longevity theory predicts that if an individual undergoes moderate stress (overheating, supercooling, decrease of caloric value of food) at the early stages of its life, this individual will live longer. However, this assumption needs careful examination in experiments. Nevertheless, it is established that
Publications:
- Kahn, Arnold, and Anders Olsen. «Stress to the rescue: Is hormesis a ‘cure’for aging?."
Dose-Response 8.1 (2010):dose-response . -
Gems, David, and Linda Partridge. «
Stress-response hormesis and aging: «that which does not kill us makes us stronger»." Cell metabolism 7.3 (2008): 200–203. - Murakami, Shin, and Thomas E. Johnson. «A genetic pathway conferring life extension and resistance to UV stress in Caenorhabditis elegans." Genetics 143.3 (1996): 1207–1218.
- Guarente, Leonard, and Cynthia Kenyon. «Genetic pathways that regulate ageing in model organisms." Nature 408.6809 (2000): 255–262.
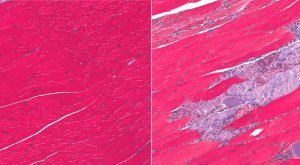 Example:
Example: The concept of phenoptosis signifies the phenomenon of programmed death of an organism.
The concept of phenoptosis signifies the phenomenon of programmed death of an organism. There are a lot of reasons why cells trigger apoptosis. If a cell finds oneself accidentally in an «alien» tissue or organ, it rapidly «commits suicide». Also, the cell infected with viruses receives a biochemical signal to make
There are a lot of reasons why cells trigger apoptosis. If a cell finds oneself accidentally in an «alien» tissue or organ, it rapidly «commits suicide». Also, the cell infected with viruses receives a biochemical signal to make  It should be noticed that evolutionary mechanisms maintaining the program of phenoptosis may be revealed. They are the kin selection (in this case, individuals age and die for the benefit of related members of a group) or the group selection (the death for the benefit of not related individuals). In theory, aging may stabilize the population, increase genetic diversity, and hasten the time of adaptation. Apoptosis in unicellular organisms (like yeasts) and the existence of organisms with «acute» programmed death (like salmon, octopus, and male marsupial mouse) are the arguments in favour of this theory.
It should be noticed that evolutionary mechanisms maintaining the program of phenoptosis may be revealed. They are the kin selection (in this case, individuals age and die for the benefit of related members of a group) or the group selection (the death for the benefit of not related individuals). In theory, aging may stabilize the population, increase genetic diversity, and hasten the time of adaptation. Apoptosis in unicellular organisms (like yeasts) and the existence of organisms with «acute» programmed death (like salmon, octopus, and male marsupial mouse) are the arguments in favour of this theory.
 Example:
Example: The dual effect of such genes is known as «antagonistic pleiotropy». Antagonistically pleiotropic genes are maintained in populations by natural selection as their early benefits outweigh their late harm.
The dual effect of such genes is known as «antagonistic pleiotropy». Antagonistically pleiotropic genes are maintained in populations by natural selection as their early benefits outweigh their late harm.
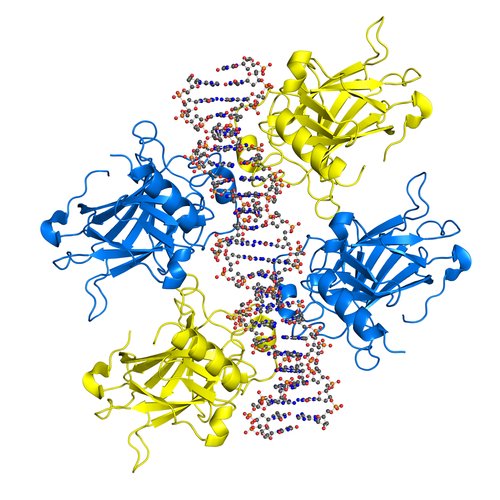 The antagonistic pleiotropy theory predicts that rapid development of an individual will correlate with rapid aging because the faster puberty is attained — the earlier senescence begins. The theory gives one more prediction: selection towards
The antagonistic pleiotropy theory predicts that rapid development of an individual will correlate with rapid aging because the faster puberty is attained — the earlier senescence begins. The theory gives one more prediction: selection towards  Existence of sexual selection in elderly age. Though the natural selection diminishes in elderly age, the evolution will promote the selection of genes important for the reproduction at an early age in rare cases only. For example, permanently changing, aggressive environment will give advantages to antagonistic pleiotropy. Nevergheless, in actual stable society where there is no abrupt changes and aggressive environment, a tendency exists to have children in later life. In this case, genes promoting healthy longevity and increasing reproductive age will be selected.
Existence of sexual selection in elderly age. Though the natural selection diminishes in elderly age, the evolution will promote the selection of genes important for the reproduction at an early age in rare cases only. For example, permanently changing, aggressive environment will give advantages to antagonistic pleiotropy. Nevergheless, in actual stable society where there is no abrupt changes and aggressive environment, a tendency exists to have children in later life. In this case, genes promoting healthy longevity and increasing reproductive age will be selected.
 History:
History: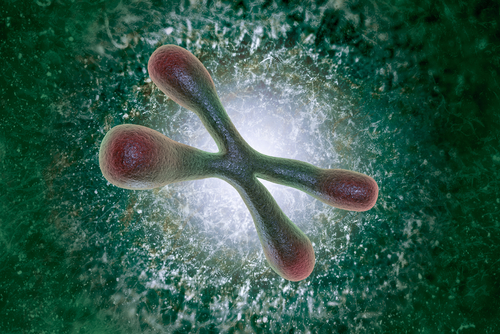 In 1961, L. Hayflick discovered that cultured human skin cells have limited capacity to divide — not more than 50 times — after which they become senescent — a phenomenon now known as the «Hayflick limit». However, Hayflick did not explain the mechanisms of this phenomenon.
In 1961, L. Hayflick discovered that cultured human skin cells have limited capacity to divide — not more than 50 times — after which they become senescent — a phenomenon now known as the «Hayflick limit». However, Hayflick did not explain the mechanisms of this phenomenon.
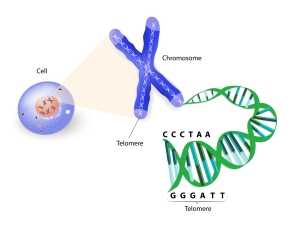 hypothesis, in each cell division, the end segments of chromosomes — telomeres — are unable to be fully copied. Therefore, telomeres become shorter after each cell division. In a certain moment, telomeres get so short, that cell becomes unable to divide. Such cell gradually lose vital capacity, and this is cell aging properly, according to the telomere theory of aging.
hypothesis, in each cell division, the end segments of chromosomes — telomeres — are unable to be fully copied. Therefore, telomeres become shorter after each cell division. In a certain moment, telomeres get so short, that cell becomes unable to divide. Such cell gradually lose vital capacity, and this is cell aging properly, according to the telomere theory of aging.
 In 1985 an enzyme called telomerase was discovered, and the Olovnikov’s theory was successfully confirmed. Telomerase maintains telomere length in cancer and germ cells, making such cells immortal. As the result, not all the cells have a limit in 50–60 divisions: cancer and germ cells have infinite replicative potential. Nevertheless, relationship of cell aging with telomeres shortening is generally acknowledged.
In 1985 an enzyme called telomerase was discovered, and the Olovnikov’s theory was successfully confirmed. Telomerase maintains telomere length in cancer and germ cells, making such cells immortal. As the result, not all the cells have a limit in 50–60 divisions: cancer and germ cells have infinite replicative potential. Nevertheless, relationship of cell aging with telomeres shortening is generally acknowledged.
 In some model organisms, such as yeast and Drosophila, there is evidence that reducing oxidative damage can extend lifespan. In mice, interventions that enhance oxidative damage generally shorten lifespan.
In some model organisms, such as yeast and Drosophila, there is evidence that reducing oxidative damage can extend lifespan. In mice, interventions that enhance oxidative damage generally shorten lifespan.
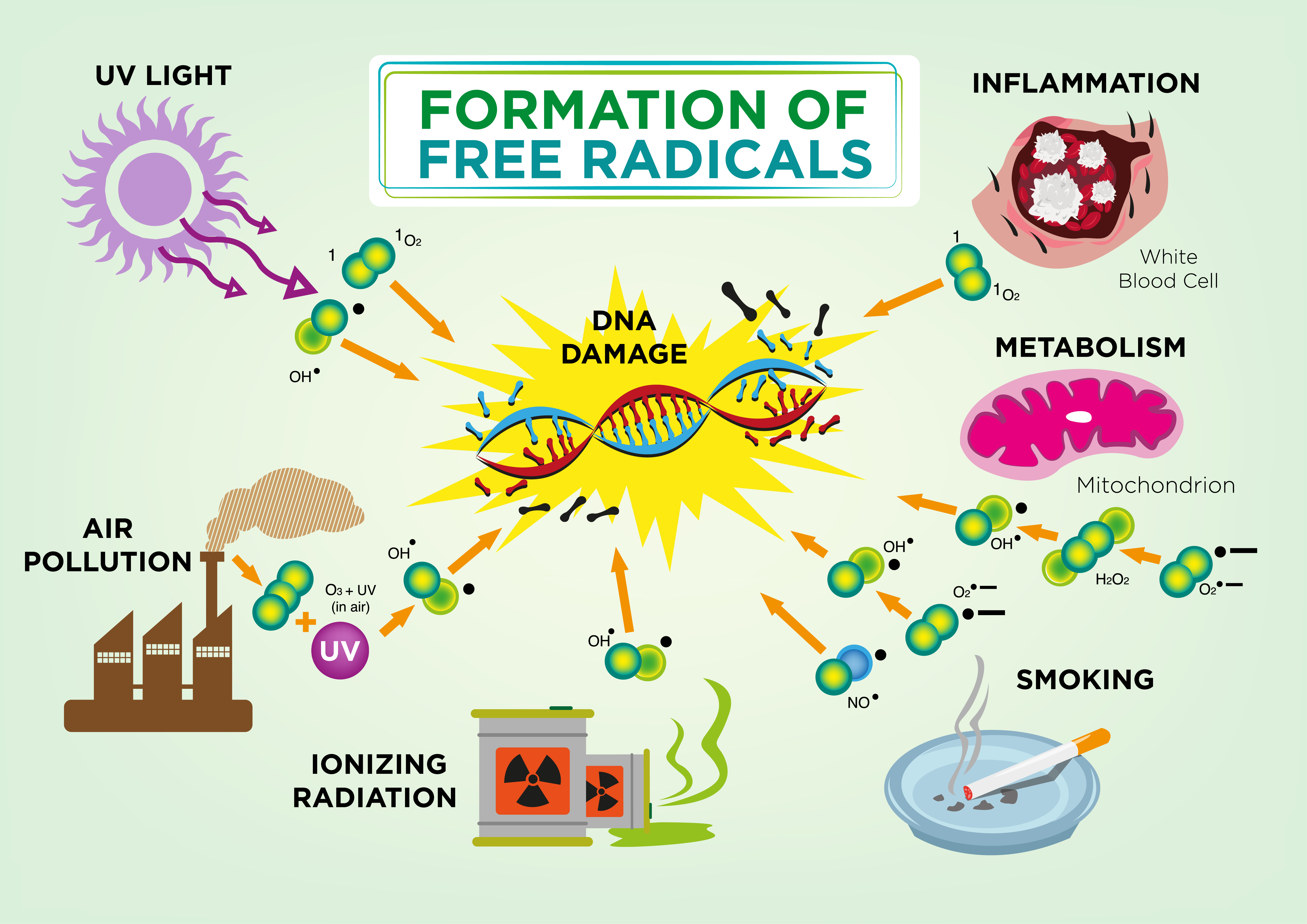 If an aggressive, chemically reactive free radical leaves that place where it is necessary, it can damage DNA, as well as RNA, proteins and lipids. Peroxidation is extremely dangerous for polyunsaturated fatty acids that are a component of cell membranes because reaction products (peroxides and hydroperoxides) has also high oxygenating potential. As the result, the process of cell damaging becomes
If an aggressive, chemically reactive free radical leaves that place where it is necessary, it can damage DNA, as well as RNA, proteins and lipids. Peroxidation is extremely dangerous for polyunsaturated fatty acids that are a component of cell membranes because reaction products (peroxides and hydroperoxides) has also high oxygenating potential. As the result, the process of cell damaging becomes  The nature provided protection mechanisms from free radicals excess, and most ROS render harmless before they injure any cell structures. The main factors of antioxidant defense of an organism are special enzymes: superoxide dismutase and some other ones. Some antioxidant chemicals are received by the organism with food. Among such antioxidants are vitamin A, vitamin C and vitamin E.
The nature provided protection mechanisms from free radicals excess, and most ROS render harmless before they injure any cell structures. The main factors of antioxidant defense of an organism are special enzymes: superoxide dismutase and some other ones. Some antioxidant chemicals are received by the organism with food. Among such antioxidants are vitamin A, vitamin C and vitamin E.
 History:
History: According to this theory, the aging of living organisms depends on casual formation of chemical bonds, or «cross links», between protein molecules. Repair enzymes of the cell can not break those bonds.
According to this theory, the aging of living organisms depends on casual formation of chemical bonds, or «cross links», between protein molecules. Repair enzymes of the cell can not break those bonds.
 The process of cross links formation between protein molecules in a human organism is very similar to the process that takes place during leather tanning. As we age, progressive accumulation of cross links occur in most tissues of our organism — in arteries, cartilages, muscles. The main consequence of this process is the decline in tissue elasticity. Actually, muscle and joint stiffness often observed in elderly people is the result of cross links formation between protein molecules.
The process of cross links formation between protein molecules in a human organism is very similar to the process that takes place during leather tanning. As we age, progressive accumulation of cross links occur in most tissues of our organism — in arteries, cartilages, muscles. The main consequence of this process is the decline in tissue elasticity. Actually, muscle and joint stiffness often observed in elderly people is the result of cross links formation between protein molecules.

 The theory was published in 1977. Actually, the disposable soma theory is the particular case of the antagonistic pleiotropy theory of aging.
The theory was published in 1977. Actually, the disposable soma theory is the particular case of the antagonistic pleiotropy theory of aging.
 This theory postulates the existence of genes which control energy resources redistribution from somatic to reproductive cells. These genes properly program the lifespan of a definite species.
This theory postulates the existence of genes which control energy resources redistribution from somatic to reproductive cells. These genes properly program the lifespan of a definite species.

 This theory receives the support from the observations on populations of wild animals in nature. These observations show that the amount and activity of predators affect the survival strategy of the population predators hunt after. For example, guppies from the population living in conditions of the increased rate of mortality grow faster and breed at earlier age than guppies living under low rate of mortality.
This theory receives the support from the observations on populations of wild animals in nature. These observations show that the amount and activity of predators affect the survival strategy of the population predators hunt after. For example, guppies from the population living in conditions of the increased rate of mortality grow faster and breed at earlier age than guppies living under low rate of mortality.

 Nowadays scientific papers present lots of circumstantial evidence supporting the mitochondrial theory of aging. There is a progbem though in studying mtDNA and mitochondria in general. Actually, scientists have only a few ways that allow working with mitochondrial mutations. There is no methods of genetic engineering suitable for mitochondria and nowadays, it is impossible to create mutation or gene, insert it into mithochondria and then study the cell line obtained. The only possible way is to study mutations given by the nature.
Nowadays scientific papers present lots of circumstantial evidence supporting the mitochondrial theory of aging. There is a progbem though in studying mtDNA and mitochondria in general. Actually, scientists have only a few ways that allow working with mitochondrial mutations. There is no methods of genetic engineering suitable for mitochondria and nowadays, it is impossible to create mutation or gene, insert it into mithochondria and then study the cell line obtained. The only possible way is to study mutations given by the nature.
 As an individual age, there is a decline in the work of neuroendocrine system. First of all, that leads to the decline in the production of many hormones that are necessary for normal functioning of an organism. Secondly, that affect hormones interractions. Althogether a lot of bodily changes connected with aging arise, e.g. menopause, decrease in muscle mass and increased chance of degenerative disease emergence and severity.
As an individual age, there is a decline in the work of neuroendocrine system. First of all, that leads to the decline in the production of many hormones that are necessary for normal functioning of an organism. Secondly, that affect hormones interractions. Althogether a lot of bodily changes connected with aging arise, e.g. menopause, decrease in muscle mass and increased chance of degenerative disease emergence and severity.

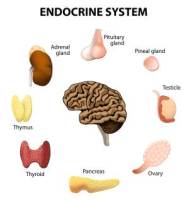
 «The main gland» of an organism is the pituitary gland. It produce a lot of hormones which control how other endocrine glands produce hormones. Hypothalamus and pituitary body control many essential functions of an organism, including internal temperature, blood pressure, thirst, hunger, sexual appetites, chemical and water balances, menstrual cycles.
«The main gland» of an organism is the pituitary gland. It produce a lot of hormones which control how other endocrine glands produce hormones. Hypothalamus and pituitary body control many essential functions of an organism, including internal temperature, blood pressure, thirst, hunger, sexual appetites, chemical and water balances, menstrual cycles.
 Initial postulates of the intoxication theory of aging were proposed by
Initial postulates of the intoxication theory of aging were proposed by  The theory postulates that aging is connected with the accumulation of different chemical agents in the cells and the organism can not use or throw this agents away.
The theory postulates that aging is connected with the accumulation of different chemical agents in the cells and the organism can not use or throw this agents away.
 Remaining side products simply accumulate in the cells. The only and universal way to fight these substances is to dilute them in the process of cell division. This method successfully works, but it works for dividing cells only. Multicellular organisms, such as human being, have problems with side products because a lot of cells stop dividing but continue to take part in the metabolism. Such cells, e.g. essential brain and heart cells, accumulate side products of the metabolism, and these products begin to impede cells' normal functioning.
Remaining side products simply accumulate in the cells. The only and universal way to fight these substances is to dilute them in the process of cell division. This method successfully works, but it works for dividing cells only. Multicellular organisms, such as human being, have problems with side products because a lot of cells stop dividing but continue to take part in the metabolism. Such cells, e.g. essential brain and heart cells, accumulate side products of the metabolism, and these products begin to impede cells' normal functioning.
 The main intracellular side product is lipofuscin, also known as
The main intracellular side product is lipofuscin, also known as  Nowadays, such scientists as B. Davidson and
Nowadays, such scientists as B. Davidson and